
Abstract
Biallelic germline mutations in MYH are associated with colorectal neoplasms, which develop through a pathway involving somatic inactivation of APC. In this study, we investigated the incidence of the common MYH mutations in an Australian cohort of sporadic colorectal cancers, the clinicopathological features of MYH cancers, and determined whether inactivation of mismatch repair and base excision repair (BER) were mutually exclusive. The MYH gene was sequenced from lymphocyte DNA of 872 colorectal cancer patients and 478 controls. Two compound heterozygotes were identified in the cancer population and all three cancers from these individuals displayed a prominent infiltration of intraepithelial lymphocytes. In total, 11 heterozygotes were found in the cancer group and five in the control group. One tumour from an individual with biallelic germline mutation of MYH also demonstrated microsatellite instability (MSI) as a result of biallelic hypermethylation of the MLH1 promoter. Although MYH-associated cancers are rare in a sporadic colorectal population, this study shows that these tumours can develop through either a chromosomal or MSI pathway. Tumours arising in the setting of BER or mismatch repair deficiency may share a biological characteristic, which promotes lymphocytic infiltration.
Similar content being viewed by others

Main
MYH-associated polyposis (MAP) is an autosomal recessive condition (Al-Tassan et al, 2002) associated with the development of multiple adenomas and carcinomas of the colon and rectum, as well as other possible malignancies and extra-colonic manifestations (Enholm et al, 2003; Sampson et al, 2003; Sieber et al, 2003; Gismondi et al, 2004; Nielsen et al, 2005). The syndrome is caused by germline biallelic mutations in MYH, a gene that codes for a DNA glycosylase involved in base excision repair (BER) (Al-Tassan et al, 2002). MYH mutations have been shown to be ethnically specific, and most (80%) Caucasian individuals with MAP possess the Y165C or G382D mutations (Enholm et al, 2003; Croitoru et al, 2004; Fleischmann et al, 2004; Wang et al, 2004; Farrington et al, 2005; Peterlongo et al, 2005). Italian populations also carry a three base pair deletion (1395delGGA) in exon 14 (Gismondi et al, 2004), whereas non-Caucasian populations harbour a range of different MYH mutations (Jones et al, 2002). As MAP is a recessive condition, patients may appear to have sporadic, rather than familial, colorectal cancer. However, a number of studies have shown that biallelic mutations in MYH account for only a minority of sporadic colorectal cancers (Enholm et al, 2003; Croitoru et al, 2004; Fleischmann et al, 2004; Wang et al, 2004; Farrington et al, 2005; Peterlongo et al, 2005). The significance of heterozygous MYH mutations in terms of cancer risk remains uncertain.
MYH-associated cancers are thought to progress through a distinct genetic pathway which does not involve microsatellite instability (MSI) (Lipton et al, 2003). Supporting this hypothesis, MYH cancers have an increased frequency of somatic transversion mutations of APC and K-ras (Al-Tassan et al, 2002; Jones et al, 2002; Lipton et al, 2003; Jones et al, 2004), although no pathologically distinctive features have been described to date.
In this study, we utilised a large and well-characterised tumour bank of sporadic colorectal cancers to investigate the clinicopathological features of MYH cancers, and to determine whether mismatch repair and base excision repair are mutually exclusive.
Materials and methods
Patients and specimens
This study was performed with the approval of the St Vincent's Human Research Ethics Committee. After obtaining informed consent, 872 individuals undergoing complete (RO or R1) surgical resection of 893 colorectal cancers at St Vincent's Hospital, Sydney, were entered in this prospective study (Ward et al, 2005). The study population consisted of 402 female and 470 male subjects with a mean age of 69.2±12.1 years (range 29–99 years). Enrolment was from 1 January 1994 to 29 May 2004, and patients presenting for resection of cancer in the setting of known inflammatory bowel disease, FAP and HNPCC were not enrolled. Peripheral blood was also collected from 284 male and 194 female healthy blood donors (Red Cross Blood Bank, Sydney), with a mean age of 45±14 years.
Tumour stage was assessed according to AJCC/UICC guidelines and histopathological characteristics were determined as previously described (Ward et al, 2005). Microsatellite status was determined using primer sets for Bat 25, Bat 26, Bat 40, D5S346, D2S123 and D17S250, and tumours with instability at two or more markers were classified as microsatellite unstable, the remainder being categorised as microsatellite stable (Ward et al, 2005). Immunohistochemistry for the mismatch repair proteins and p53 was performed as previously described (Ward et al, 2005).
MYH mutation analysis
To identify the common ‘Caucasian’ pathogenic MYH mutations, exons 7, 13 and 14 of MYH were amplified and sequenced from lymphocyte DNA using previously designed primers (Gismondi et al, 2004; Kairupan et al, 2005). The Y165C (exon 7) and 1395delGGA (exon 14) mutations were confirmed by sequencing the complementary strand, and the G382D (exon 13) mutation was verified using a restriction enzyme digest (BglII). The remaining MYH exons were then sequenced in those individuals who were heterozygotes for one of the common mutations. Briefly, exon-specific primers ((Gismondi et al, 2004; Kairupan et al, 2005) additional details available on request) were used to amplify the 16 exons of MYH in 25 μl reactions containing 100 ng of genomic DNA, 0.4 μ M of each of the primer, 0.2 μ M of dNTP, 1 U of FastStart Taq polymerase and a PCR master mix at 1.5 mM MgCl2. The PCR products were purified using the QIAquick PCR purification kit (Qiagen, Basel, Switzerland) and sequencing was performed using the Big Dye Terminator Cycle Sequencing kit (Applied Biosystems, Rotkreuz, Switzerland). Purified sequence reactions were separated on the ABI3100 Genetic Analyser (Applied Biosystems), and the nucleotide sequences analysed with the aid of the Sequencher software program (Gene Codes Corporation, Ann Arbor, MI, USA).
Methylation analysis of MLH1 promoter
DNA (1–2 μg) was converted with sodium bisulphite and the ‘A’ and ‘C’ regions of the MLH1 promoter were subject to combined bisulphite restriction analyses (COBRA) as previously described (Hitchins et al, 2005). Samples giving a positive restriction digestion pattern indicative of the presence of methylation were cloned into the pGEMTeasy vector (Promega, Annandale, NSW, Australia), and individual clones were fluorescently sequenced on an ABI 3100 using SP6 and T7 vector primers.
Statistical analysis
Categorical variables were compared using the χ2-test or Fisher's exact test as appropriate. In patients with more than one cancer, only the properties of the most advanced tumour were considered in the case control study. All tumours were considered when analysing tumour properties. A probability value of less than 0.05 was considered significant. All data were analysed using the SPSS statistical software V13.0 (SPSS Inc., Chicago, IL, USA).
Results
Frequency and spectrum of MYH mutations
Of the 872 individuals with colorectal cancer, two were compound heterozygotes (patient 9033 and 13714) and a further 11 had a monoallelic mutation in MYH (Table 1). None of the 478 control individuals harboured biallelic MYH mutations, but five were found to be heterozygotes (Table 1). After sequencing the remaining exons in the 16 heterozygotes, a number of nonpathogenic missense mutations were identified (one V22M, five Q324H and one S501F). No significant difference in the frequency of biallelic (P=0.542, Fisher's exact test) or monoallelic MYH mutations (χ2=0.12, P=0.72) was found between the colorectal and the control groups.
Pathological features of tumours in MYH mutation carriers
At the time of surgical resection, one compound heterozygote (patient 9033) was found to have synchronous stage III colorectal cancers as well as more than 100 adenomatous polyps scattered throughout the colon. The clinicopathological characteristics of these cancers, as well as the stage III caecal cancer from individual 13714 (G382D/1395delGGA) are shown in Table 2. Both of these individuals harboured multiple adenomatous polyps; however the only manifestation of extra-colonic disease was in patient 9033, who had a history of malignant melanoma at the age of 28 years. Interestingly, the three cancers from these individuals displayed a prominent infiltration of intraepithelial lymphocytes.
In most respects, cancers arising in monoallelic MYH carriers were indistinguishable from cancers arising in individuals in whom no MYH mutations had been identified (Table 2). A statistically significant increase in the number of intraepithelial lymphocytes was noted in the heterozygote mutation group compared with the rest of the colorectal cohort (five of 11 vs 151 of 827 assessable cases, P=0.04, Fisher's Exact test). However, this association was probably a reflection of the microsatellite status of the cancer, as all but one case with prominent intraepithelial lymphocytes was associated with a microsatellite unstable cancer.
Microsatellite instability and MYH mutations
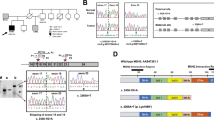
The frequency of tumours with microsatellite instability arising in cases with MYH mutations is shown in Table 2. Of the 893 tumours assessed for MSI in this study, 139 (15.5%) showed MSI. Individuals who were heterozygous for an MYH mutation were more likely to have an MSI tumour (five of 11, χ2 test P<0.01), whereas one of the cancers seen in the two individuals with biallelic MYH mutations was also MSI (Table 2). This individual (9033) had a microsatellite unstable caecal cancer which failed to express the mismatch repair protein MLH1 by immunohistochemistry, as well as a rectal cancer that was microsatellite stable and expressed the mismatch repair proteins MLH1, MSH2 and MSH6. The microsatellite unstable caecal cancer did not display the typical pathological features of a sporadic MSI colorectal cancer, in that it was nonmucinous, of low histological grade and BRAF mutation negative (data not shown). The pedigree and results of MYH mutation testing in this family is shown in Figure 1, panel A. Germline testing of the mismatch repair genes failed to identify a pathogenic mutation in individual 9033.

(A) Pedigree showing the family of the proband (case 9033, marked with black arrow) and their respective disease and mutation status. WT, wild type; left half shaded black, polyps; right half shaded black, cancer. (B) Schematic of promoter region of MLH1 from the caecal tumour from case 9033 demonstrating biallelic methylation. Each allele is distinguished by the presence of an SNP (coloured square). Circles represent CpG islands, filled circles represent methylation, the blue square represents guanine and the red square represents adenine. (C) Sequence traces obtained from the caecal tumour of case 9033 demonstrate the presence of biallelic MYH mutations. The patient was compound heterozygote for the Y165C and G382D mutations.
To determine the mechanism of inactivation of MLH1 in the caecal cancer of patient 9033, we subjected the MLH1 promoter to bisulphite sequencing. As patient 9033 was heterozygous for the A/G polymorphism (rs11800734) in the MLH1 promoter, bisulphite sequencing was able to confirm methylation of both alleles in the tumour cells (Figure 1B). Unmethylated A and G alleles were presumably derived from contaminating normal tissue, thus suggesting that normal somatic colonic cells were unmethylated. Microdissected tumour tissue was then subjected to genetic analysis for MYH mutations. As expected, the tumour displayed the missense mutations Y165C and G382D (Figure 1C), thus confirming that this carcinoma possessed MSI in the context of biallelic MYH mutations.
Discussion
This study shows that biallelic MYH mutations are rare (0.2%, 2/872) in an Australian colorectal cancer population. It must be acknowledged that the screening strategy used in this study examined exons 7, 13 and 14 of MYH. While these regions account for around 80% of known mutations in Caucasians, a small number of pathogenic mutations will have been missed because of this approach. Nevertheless, this mutation frequency was comparable to studies from Finland (0.4%, four out of 1042) (Enholm et al, 2003), North America (0.4%, two out of 555 and two out of 444, 0.5%) (Wang et al, 2004; Peterlongo et al, 2005), Canada (1.0%, 12 out of 1238) (Croitoru et al, 2004), Scotland (0.5%, 12/2239)(Farrington et al, 2005) and Britain (0.3%, one out of 358) (Fleischmann et al, 2004). Like other studies, we found a lower frequency of the Y165C allele (0.2% in cases and 0.1% in controls) than the G382D allele (0.6% in cases and 0.4% in controls). Given the significant number of southern European migrants resident in Australia, it is surprising that only one individual was identified with an exon 14 mutation (135delGGA) (Gismondi et al, 2004). While others have suggested that the frequency mutations in exon 14 such as A459T justify routine mutation screening of this exon, our data does not support this position, at least in a nonpolyposis population (Alhopuro et al, 2005).
Lipton et al (2003) proposed that carcinomas arising in the setting of biallelic MYH mutations followed a distinct genetic pathway. If this hypothesis was correct, a distinctive clinicopathological phenotype may accompany the development of MYH-related cancers. Although we found little evidence to support this suggestion, it was interesting that the three cancers arising in the individuals with biallelic mutations in MYH displayed a prominent infiltration of intraepithelial lymphocytes. Lymphocytic infiltrates are strongly associated with the development of mismatch repair-deficient cancers, although a proportion of microsatellite stable tumours also demonstrate intraepithelial lymphocytes (Ward et al, 2001). The mechanism for recruitment of lymphocytes in these cancers is unknown (Quinn et al, 2003), but mutator and DNA repair phenotypes may share properties which favour the retention of lymphocytes in the epithelium. The phenomenon of increased lymphocytic infiltration in MSI tumours has been suggested as a possible reason for the improved survival seen in these cancers (Guidoboni et al, 2001). It is possible that MAP cancers represent a subgroup of MSS cancers that may also share this improved prognosis.
Significantly, our study has also shown that MSI cancers can arise in the setting of biallelic germline MYH mutations, and that MSI cancers may arise more commonly in individuals with monoallelic MYH germline mutation than in normal individuals. To date, the 168 MAP tumours (includes 48 colorectal cancers) reported in the literature were microsatellite stable (Lipton et al, 2003; Fleischmann et al, 2004; Wang et al, 2004; Nielsen et al, 2005). This is expected given that mutations of MYH predisposes to inactivation of APC through G:C to A:T transversions (Al-Tassan et al, 2002). The absence of MSI in MAP carcinomas led to the suggestion that the derangement of both base excision repair and mismatch repair was not compatible with cellular survival (Kambara et al, 2004). The case of an MSI cancer in the setting of MYH biallelic germline mutation reported in our study clearly does not support this hypothesis, and it is interesting that MLH1 was inactivated by biallelic promoter methylation, rather than through somatic mutations secondary to loss of MYH function. In this case, it is apparent that biallelic MYH mutations have played a role in the early stages of colorectal carcinogenesis through APC mutation and polyp formation. Yet, this influence has been superseded by the tumorigenic drive provided by loss of MLH1 function. It is possible that MYH mutations may have a largely permissive role in tumorigenesis through polyp formation, with subsequent progression occurring as a result of changes to other key genes. This role of MYH mutations in colorectal carcinogenesis is consistent with the finding in murine models that MYH knockouts are phenotypically normal (Xie et al, 2004).
It is also worth noting that patient 9033 was only 50 years of age. This is a strikingly young age for the development of a ‘sporadic’ MSI cancer through biallelic methylation of MLH1 (Ward et al, 2001), and raises the possibility that MYH mutation has somehow either facilitated methylation, or acted synergistically with MLH1 loss to more rapidly drive tumour progression. A more testable implication is that MYH mutation may be worth considering in younger individuals with MSI tumours that have developed because of MLH1 methylation. Such individuals are increasingly likely to be recognised because of immunohistochemical testing of colorectal cancers. Biallelic MYH mutation remains an uncommon event, and the small numbers of cases in this report allow us only to speculate on the biological relevance in a sporadic population. More systematic studies of MYH status in distinct populations will be required to properly evaluate the true significance of MYH loss in colorectal carcinogenesis.
Change history
16 November 2011
This paper was modified 12 months after initial publication to switch to Creative Commons licence terms, as noted at publication
References
Alhopuro P, Parker AR, Lehtonen R, Enholm S, Jarvinen HJ, Mecklin JP, Karhu A, Eshleman JR, Aaltonen LA (2005) A novel functionally deficient MYH variant in individuals with colorectal adenomatous polyposis. Hum Mutat 26: 393
Al-Tassan N, Chmiel NH, Maynard J, Fleming N, Livingston AL, Williams GT, Hodges AK, Davies DR, David SS, Sampson JR, Cheadle JP (2002) Inherited variants of MYH associated with somatic G : C–>T : A mutations in colorectal tumors. Nat Genet 30: 227–232
Croitoru ME, Cleary SP, Di Nicola N, Manno M, Selander T, Aronson M, Redston M, Cotterchio M, Knight J, Gryfe R, Gallinger S (2004) Association between biallelic and monoallelic germline MYH gene mutations and colorectal cancer risk. J Natl Cancer Inst 96: 1631–1634
Enholm S, Hienonen T, Suomalainen A, Lipton L, Tomlinson I, Karja V, Eskelinen M, Mecklin JP, Karhu A, Jarvinen HJ, Aaltonen LA (2003) Proportion and phenotype of MYH-associated colorectal neoplasia in a population-based series of Finnish colorectal cancer patients. Am J Pathol 163: 827–832
Farrington SM, Tenesa A, Barnetson R, Wiltshire A, Prendergast J, Porteous M, Campbell H, Dunlop MG (2005) Germline susceptibility to colorectal cancer due to base-excision repair gene defects. Am J Hum Genet 77: 112–119
Fleischmann C, Peto J, Cheadle J, Shah B, Sampson J, Houlston RS (2004) Comprehensive analysis of the contribution of germline MYH variation to early-onset colorectal cancer. Int J Cancer 109: 554–558
Gismondi V, Meta M, Bonelli L, Radice P, Sala P, Bertario L, Viel A, Fornasarig M, Arrigoni A, Gentile M, Ponz de Leon M, Anselmi L, Mareni C, Bruzzi P, Varesco L (2004) Prevalence of the Y165C, G382D and 1395delGGA germline mutations of the MYH gene in Italian patients with adenomatous polyposis coli and colorectal adenomas. Int J Cancer 109: 680–684
Guidoboni M, Gafa R, Viel A, Doglioni C, Russo A, Santini A, Del Tin L, Macri E, Lanza G, Boiocchi M, Dolcetti R (2001) Microsatellite instability and high content of activated cytotoxic lymphocytes identify colon cancer patients with a favorable prognosis. Am J Pathol 159: 297–304
Hitchins M, Williams R, Cheong K, Halani N, Lin VA, Packham D, Ku S, Buckle A, Hawkins N, Burn J, Gallinger S, Goldblatt J, Kirk J, Tomlinson I, Scott R, Spigelman A, Suter C, Martin D, Suthers G, Ward R (2005) MLH1 germline epimutations as a factor in hereditary nonpolyposis colorectal cancer. Gastroenterology 129: 1392–1399
Jones S, Emmerson P, Maynard J, Best JM, Jordan S, Williams GT, Sampson JR, Cheadle JP (2002) Biallelic germline mutations in MYH predispose to multiple colorectal adenoma and somatic G : C–>T : A mutations. Hum Mol Genet 11: 2961–2967
Jones S, Lambert S, Williams GT, Best JM, Sampson JR, Cheadle JP (2004) Increased frequency of the k-ras G12C mutation in MYH polyposis colorectal adenomas. Br J Cancer 90: 1591–1593
Kairupan CF, Meldrum CJ, Crooks R, Milward EA, Spigelman AD, Burgess B, Groombridge C, Kirk J, Tucker K, Ward R, Williams R, Scott RJ (2005) Mutation analysis of the MYH gene in an Australian series of colorectal polyposis patients with or without germline APC mutations. Int J Cancer 116: 73–77
Kambara T, Whitehall VL, Spring KJ, Barker MA, Arnold S, Wynter CV, Matsubara N, Tanaka N, Young JP, Leggett BA, Jass JR (2004) Role of inherited defects of MYH in the development of sporadic colorectal cancer. Genes Chromosomes Cancer 40: 1–9
Lipton L, Halford SE, Johnson V, Novelli MR, Jones A, Cummings C, Barclay E, Sieber O, Sadat A, Bisgaard ML, Hodgson SV, Aaltonen LA, Thomas HJ, Tomlinson IP (2003) Carcinogenesis in MYH-associated polyposis follows a distinct genetic pathway. Cancer Res 63: 7595–7599
Nielsen M, Franken PF, Reinards TH, Weiss MM, Wagner A, van der Klift H, Kloosterman S, Houwing-Duistermaat JJ, Aalfs CM, Ausems MG, Brocker-Vriends AH, Gomez Garcia EB, Hoogerbrugge N, Menko FH, Sijmons RH, Verhoef S, Kuipers EJ, Morreau H, Breuning MH, Tops CM, Wijnen JT, Vasen HF, Fodde R, Hes FJ (2005) Multiplicity in polyp count and extracolonic manifestations in 40 Dutch patients with MYH associated polyposis coli (MAP). J Med Genet 42: e54
Peterlongo P, Mitra N, Chuai S, Kirchhoff T, Palmer C, Huang H, Nafa K, Offit K, Ellis NA (2005) Colorectal cancer risk in individuals with biallelic or monoallelic mutations of MYH. Int J Cancer 114: 505–507
Quinn E, Hawkins N, Yip YL, Suter C, Ward R (2003) CD103+ intraepithelial lymphocytes – a unique population in microsatellite unstable sporadic colorectal cancer. Eur J Cancer 39: 469–475
Sampson JR, Dolwani S, Jones S, Eccles D, Ellis A, Evans DG, Frayling I, Jordan S, Maher ER, Mak T, Maynard J, Pigatto F, Shaw J, Cheadle JP (2003) Autosomal recessive colorectal adenomatous polyposis due to inherited mutations of MYH. Lancet 362: 39–41
Sieber OM, Lipton L, Crabtree M, Heinimann K, Fidalgo P, Phillips RK, Bisgaard ML, Orntoft TF, Aaltonen LA, Hodgson SV, Thomas HJ, Tomlinson IP (2003) Multiple colorectal adenomas, classic adenomatous polyposis, and germ-line mutations in MYH. N Engl J Med 348: 791–799
Wang L, Baudhuin LM, Boardman LA, Steenblock KJ, Petersen GM, Halling KC, French AJ, Johnson RA, Burgart LJ, Rabe K, Lindor NM, Thibodeau SN (2004) MYH mutations in patients with attenuated and classic polyposis and with young-onset colorectal cancer without polyps. Gastroenterology 127: 9–16
Ward R, Meagher A, Tomlinson I, O'Connor T, Norrie M, Wu R, Hawkins N (2001) Microsatellite instability and the clinicopathological features of sporadic colorectal cancer. Gut 48: 821–829
Ward RL, Turner J, Williams R, Pekarsky B, Packham D, Velickovic M, Meagher A, O'Connor T, Hawkins NJ (2005) Routine testing for mismatch repair deficiency in sporadic colorectal cancer is justified. J Pathol 207: 377–384
Xie Y, Yang H, Cunanan C, Okamoto K, Shibata D, Pan J, Barnes DE, Lindahl T, McIlhatton M, Fishel R, Miller JH (2004) Deficiencies in mouse Myh and Ogg1 result in tumor predisposition and G to T mutations in codon 12 of the K-ras oncogene in lung tumors. Cancer Res 64: 3096–3102
Acknowledgements
This study was supported by NSW Cancer Council and National Health and Medical Research Council of Australia. AC was supported by a scholarship from the Royal College of Pathologists of Australasia.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
From twelve months after its original publication, this work is licensed under the Creative Commons Attribution-NonCommercial-Share Alike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Colebatch, A., Hitchins, M., Williams, R. et al. The role of MYH and microsatellite instability in the development of sporadic colorectal cancer. Br J Cancer 95, 1239–1243 (2006). https://doi.org/10.1038/sj.bjc.6603421
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.bjc.6603421
Keywords
This article is cited by
-
Recent Advances on Immune Targeted Therapy of Colorectal Cancer Using bi-Specific Antibodies and Therapeutic Vaccines
Biological Procedures Online (2021)
-
Role of DNA repair defects in predicting immunotherapy response
Biomarker Research (2020)
-
Association of monoallelic MUTYH mutation among Egyptian patients with colorectal cancer
Familial Cancer (2017)
-
Germline variants in POLE are associated with early onset mismatch repair deficient colorectal cancer
European Journal of Human Genetics (2015)
-
Biallelic MUTYH mutations can mimic Lynch syndrome
European Journal of Human Genetics (2014)
