Article Text

Abstract
The gut microbiota is known to play an important role in maintaining gut health through a symbiotic relationship with the host. Altered gut microbiota is a common feature of several diseases of the gastrointestinal tract; however, the causal relationship between microbiota and disease pathogenesis is poorly understood. Necrotising enterocolitis (NEC) and inflammatory bowel disease (IBD) are both severe inflammatory diseases affecting the gastrointestinal tract. Although they affect very different patient populations, with NEC primarily being a disease of prematurity and IBD predominantly affecting adults although children can be affected, they both demonstrate common features of gut microbial dysbiosis and a dysregulated host immune response. By comparing and contrasting the changes in gut microbiota, host immune response and function, we aim to highlight common features in diseases that may seem clinically unrelated. Key areas of interest are the role of pattern recognition receptors in altered recognition and responses to the gut microbiota by the host immune system and the associated dysfunctional gut epithelial barrier. The challenge of identifying causal relationships between microbiota and disease is ever-present; however, considering a disease-agnostic approach may help to identify mechanistic pathways shared across several clinical diseases.
- NECROTIZING ENTEROCOLITIS
- INFLAMMATORY BOWEL DISEASE
- IBD
- INTESTINAL BACTERIA
Statistics from Altmetric.com
Introduction
Approximately 100 trillion (1×1014) microbes colonise the human gastrointestinal (GI) tract, composed of bacteria, fungi, yeasts, viruses, protozoa and archaea.1 2 Over 1000 distinct commensal bacterial species from four dominant phyla: Firmicutes, Bacteroidetes, Actinobacteria and Proteobacteria 3 are found along the entire length of the GI tract, with increasingly dense and varied populations in the colon.4 These complex microbial communities have developed a symbiotic relationship with the human host that is the result of over 500 million years of coevolution.5
The genes and genetic products of the microbiota are collectively referred to as the gut microbiome and they are essential for normal gut function and homoeostasis. With the use of culture-independent sequencing techniques (16S rRNA and shotgun metagenomics) the composition and function of the human gut microbiome is being uncovered. International projects, such as The Human Microbiome Project, have been undertaken to characterise the commensal microbiota in distinct anatomical sites of healthy individuals.3 Although these are still mostly focused on European and North American populations,6 this has provided better understanding of the complex and varied roles the gut microbiota plays in both health and disease.
This review will consider how the gut microbiota develops from early life onwards, its key functions in maintaining gut health and the role of altered gut microbiota in disease. Although altered gut microbiota is a characteristic feature of several diseases, whether there is a causal link remains poorly understood.
Necrotising enterocolitis (NEC) and inflammatory bowel disease (IBD) are both severe inflammatory diseases of the GI tract. They affect very different patient populations, with NEC being a disease of prematurity and IBD primarily affecting those in the second and third decades of life. Although host-specific factors differ in these patient groups, both conditions are associated with distinct changes to the composition of the gut microbiome, and it is postulated that these might drive aberrant function of the gut immune system resulting in destructive inflammation. We describe the role of the gut microbiota in the pathogenesis of these conditions and draw parallels and differences between them. By highlighting features of the immature gut immune system characteristic of those with NEC and those of the genetically susceptible host in IBD we will consider the potential causal role for host–microbe interactions in promoting gut inflammation.
Development of the gut microbiota
Age greatly influences the composition and diversity of the gut microbiota, with key stages throughout life driving substantial shifts in bacterial abundances.7 8 Of these changes, early life factors have a profound impact, with mode of delivery, feeding type and then transitioning to a solid based diet driving alterations in microbiota composition.9 As such, the assembly and trajectory of the gut microbiota is not purely stochastic, but instead follows a patterned development.10 11
The sequential changes in the initial neonatal gut microbiota composition are well defined and follow a predictable, non-random sequence for healthy, full-term neonates (figure 1).12 13 Initially, the gut is colonised by facultative anaerobes belonging to the Proteobacteria phylum, predominantly the Enterobacteriaceae family of bacteria, which are believed to deoxygenate the gut.9 14 Once an anaerobic environment is formed in the gut, obligate anaerobes begin to colonise, including Bifidobacterium which belong to the Actinobacteria phylum.13–15
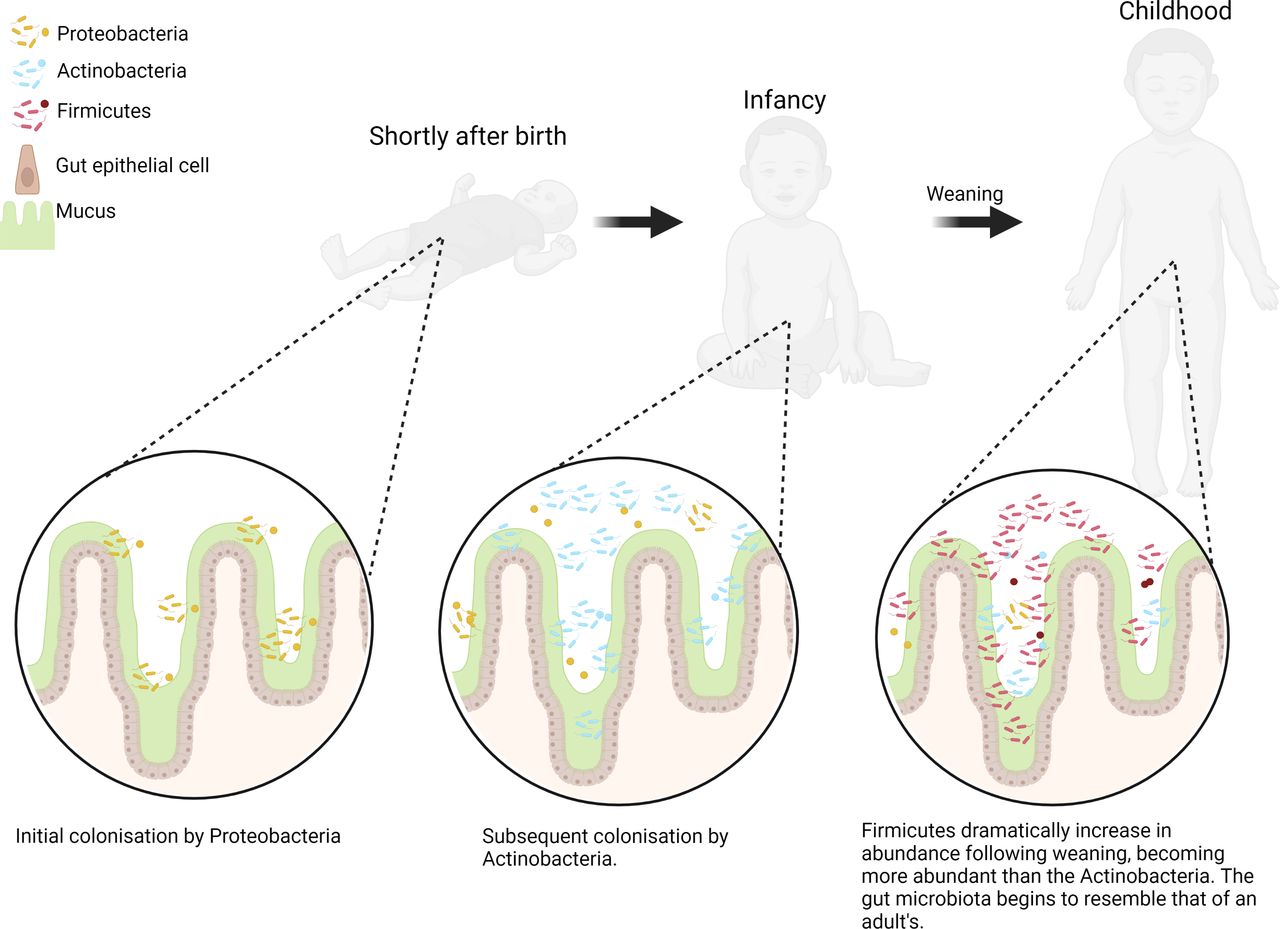
The early development of the gut microbiota. Key changes to the gut microbiota composition are summarised on a phylum level with changes shown at three early life stages: shortly after birth, infancy and postweaning (figure created in BioRender).
Transitioning to a solid based diet from milk is an important milestone in the microbiota development of the neonate and denotes a shift towards a microbiota which resembles that of an adult’s.7 12 16 17 A cross-sectional study of the gut microbiota from new-borns to centenarians shows a dramatic change at 2 years of age whereby the Firmicutes dramatically increase in abundance with the Actinobacteria declining, likely corresponding to the period following weaning.7 Comparisons of the gut microbiota before and after weaning in two distinct geographical cohorts also shows some clear and dramatic changes.16
Much research has been conducted to understand the factors contributing to interindividual variability seen in gut microbiota compositions of adults. In particular, the effect of diet and geography have been studied extensively.18–22 Diets are broadly divided into Western defined diets, comprising relatively lower levels of fibre and higher consumption of refined carbohydrates and fat, and high fibre or Eastern diets.21 The diversity of the gut microbiota community in Western countries is significantly lower than that of non-Western ones with diet thought to be an important determinant of diversity.23
There is a complex symbiotic relationship between the resident microbiota and the host, with a diverse gut microbiota being necessary for normal gut function and health.
Role of the microbiota in normal gut function and protection against pathogens
The organisms comprising the gut microbiota have a symbiotic relationship with the human host and perform several essential functions including regulating host metabolism, synthesis of essential vitamins and protection against pathogens.24 25 In return, these organisms can occupy a nutrient rich environment. The gut microbiota plays an essential role in host digestive function and metabolism, by enabling the extraction of nutrients from substrates that are otherwise indigestible by the host.26 This is primarily achieved through fermentation and anaerobic degradation to maximise the caloric uptake from these ingested nutrients. Metabolites produced through this breakdown of insoluble carbohydrates have a variety of functions in gut homoeostasis. Short chain fatty acids (SCFAs) are the most abundant of these metabolites and include acetate, propionate and butyrate.27 These SCFAs are either used locally as a fuel source by the colonic intestinal epithelial cells (IECs) or absorbed into the portal circulation.28
There are several commensal butyrate-producing species in the colon, with Firmicutes (eg, Faecalibacterium prausnitzii) and Bacteroidetes being the predominant groups.29 Butyrate provides an essential source of energy to the colonic mucosal epithelium, stimulating growth and proliferation of IECs and having an anti-inflammatory effect (table 1).30 31 This anti-inflammatory effect is achieved via several mechanisms including enhancement of intestinal barrier function by increasing mucin production and maintaining the integrity of epithelial tight junctions. Butyrate also reduces expression of inflammatory mediators by IECs via inhibition of the proinflammatory NF-kB pathway.32 33
Role of butyrate in intestinal homoeostasis
Gut microbial dysbiosis and gut inflammation: a causal relationship?
Dysbiosis can be defined as a compositional and functional alteration in the microbiota in individuals with disease compared with healthy subjects.34 This is a feature of several intestinal and extra-intestinal disorders including IBD and NEC, colorectal cancer, diabetes, multiple sclerosis, obesity and food allergies.25 ,35–37
NEC is an acute and devastating inflammatory disease of the intestine which leads to gut tissue necrosis primarily affecting preterm infants.38 39 Risk factors include low gestational age and birth weight, formula milk feeding and administration of antibiotics, with varying degrees of risk.40–43
The involvement of gut bacteria in NEC pathogenesis is apparent as the disease is associated with altered gut microbiota in several animal models and human cases of NEC (table 2).39 44–50 Several studies have analysed the relative abundance of bacterial taxa preceding NEC, using a 16S rRNA sequencing approach. A meta-analysis of 14 eligible studies, either cohort or case–control, has found that the phylum Proteobacteria appear to have an increased relative abundance in NEC infants compared with healthy non-NEC control infants.51 Whether this increase in Proteobacteria abundance preceding NEC onset is causative is unknown. It may be in response to other factors which are drivers of NEC such as hypoxia in the gut environment.
A comparison of the major microbial changes seen in the gut microbiotas for IBD and NEC
IBD, largely comprised of Crohn’s disease (CD) and ulcerative colitis (UC), is another condition associated with inflammation of the GI tract. IBD is associated with specific alterations in the gut microbiota (table 2). Changes frequently reproduced across several studies are that of reduced richness and diversity of both the faecal and mucus-resident microbiome in IBD.52–54 Specifically, reduction in relative abundance of Faecalibacterium prausnitzii and an increase in members of the Proteobacteria phylum such as Escherichia coli are observed in people with IBD compared with healthy individuals.55 56 These differences are seen among members of the same family, including twins, who are discordant for IBD implying that alterations in the gut microbiota are primarily related to disease state.57
Reduced bacterial diversity and shifts in abundance of specific bacterial taxa have also been associated with several chronic inflammatory pathologies, including other immune-mediated conditions such as type 1 diabetes.58 59 The commonality in these findings questions the specificity of dysbiosis in defining disease states. In particular, whether the observed alterations in the microbiota are a cause or consequence of inflammation is yet to be clearly defined.
Role of pattern recognition receptors in promoting gut inflammation
While clear cause and correlation has not been established, there are several mechanisms by which an altered gut microbiota can influence the onset of gut inflammation (figure 2). One important way is by altering the response to recognition of commensal microbes by the host immune system. Genome-wide association studies have identified a variety of target genes implicated in susceptibility to IBD.60 These susceptibility loci and mutations are associated with defects in microbial sensing and clearance.

How pattern recognition receptors expressed on intestinal epithelial cells (IECs) may be involved in necrotising enterocolitis (NEC) and inflammatory bowel disease (IBD) pathology. Alterations to the function and expression of TLR4 on intestinal epithelial cells may beimportant in contributing to the gut microbiota dysbiosis seen during both inflammatorybowel disease and necrotising enterocolitis and subsequently disease pathology. NOD2receptor mutations have also been implicated in IBD pathology via altering the productionof AMPs which may alter the gut microbiota composition leading to dysbiosis and gutinflammation. These are putative mechanisms as indicated by ?. TLR4- Toll-like receptor 4,IECs-Intestinal epithelial cells, AMPs-Antimicrobial peptides, NOD2-Nucleotide-bindingoligomerization domain-containing protein 2. (Figure created in BioRender).
Pattern recognition receptors (PRRs) are integral to the interaction between the host and gut microbiota. They are expressed on IECs and many immune cells and recognise pathogen-associated molecular patterns found in microbes. PRRs are divided into four main families: Toll-like receptors (TLR), nucleotide-binding oligomerisation domain-like receptors (NLR), C-type lectin receptors and RIG-1 like receptors. Altered function and expression of TLRs and NLRs have been specifically implicated in the pathogenesis of IBD and NEC, two conditions linked with microbial dysbiosis.
The role of PRRs, specifically TLR4 have been extensively studied in the pathogenesis of NEC (figure 2).61 TLR4, present on the surface of IECs, recognises and binds to bacterial LPS, which is a component of the outer surface of gram-negative organisms. The binding of LPS to TLR4 initiates the NF-kB signalling pathway leading to the production of inflammatory cytokines.62 Observations that there is increased TLR4 expression on IECs in the foetal gut compared with the full-term neonate have led researchers to consider the role of TLR4 in the aetiology of NEC.63 The reasons for this temporal alteration in TLR4 expression remains unclear, however, TLR4 downregulation may be a mechanism to promote tolerance of gram negative organisms when colonised on birth.63–65
Diet and gut hypoxia are also important in altered gut TLR4 expression. Rat and mouse models of NEC have demonstrated that intestinal TLR4 expression is significantly increased for formula fed hypoxia-stressed litters relative to those which are breastfed and are not hypoxia stressed.46 66 Human infants who develop clinical NEC also show increased intestinal TLR4 expression compared with those who do not.66 This relative increase in TLR4 expression may be related to the immature status of the preterm infant gut and immune system.
Although the aetiology of NEC is poorly defined, it is hypothesised that the immature gut and immune system interact with an altered gut microbiota. This leads to excessive and damaging gut inflammation. Preceding the onset of NEC, there is an increased abundance of gram-negative organisms such as Proteobacteria. These organisms contain LPS in their outer walls, leading to excessive gut inflammation via the LPS-TLR4 interaction.51 67 68
TLR4 polymorphisms have also been shown to be associated with susceptibility to both CD and UC.69 Furthermore, there is increased expression of TLR4 by IECs in both adult and paediatric populations with IBD (figure 2).70 Several studies have demonstrated an increased abundance of Enterobacteriaceae, specifically adherent-invasive E. coli found in tissue biopsies of people with CD.71 72 These organisms contain LPS in their outer walls, suggesting a role for LPS-TLR4 interaction driven inflammation similar to that previously described in NEC.
Another PRR linked to gut inflammation is NOD2. NOD2/CARD15 (nucleotide-binding oligomerisation domain 2) is one of a family of intracellular PRRs which recognise components from the degradation of microbial cell walls such as muramyl dipeptides. Mutations of NOD2, leading to decreased responsiveness to its ligands is associated with for example reduced production of antibacterial peptides, and has been consistently linked with susceptibility to CD and even increased risk of colorectal cancer (figure 3).73–77 NOD2 deficient mice demonstrate a specific mucosa-associated microbial dysbiosis with a higher proportion of Bacteroidetes, lower representation of Firmicutes in the colon and an increase in the relative abundance of pathogenic gram-negative bacterial species.78 In chemically induced mouse models of colitis, this NOD2-mediated dysbiosis can increase risk of inflammation with this risk being transmissible to wild-type mice colonised by faecal transfer with the same dysbiotic microbiota.79 Such changes in the microbiota are mirrored in human studies of CD,52 ,80 with a decrease in overall species richness, alpha-diversity and depletion of commensal species of the phyla Firmicutes and increased abundance of E. coli.56 Alterations in NOD2 signalling have also been implicated in driving inflammatory responses by promoting the production of inflammatory cytokines thus propagating inflammation. To our knowledge, the role of NOD2 has not been assessed in NEC but alterations in the function of several PRR may contribute to both conditions (figure 3). Furthermore, it is notable that there is cooperation between PRR signalling pathways including NOD2, TLR3, 4 and 9 to enhance inflammatory responses.81–83
{kind=link}
{kind=link}
{kind=link}
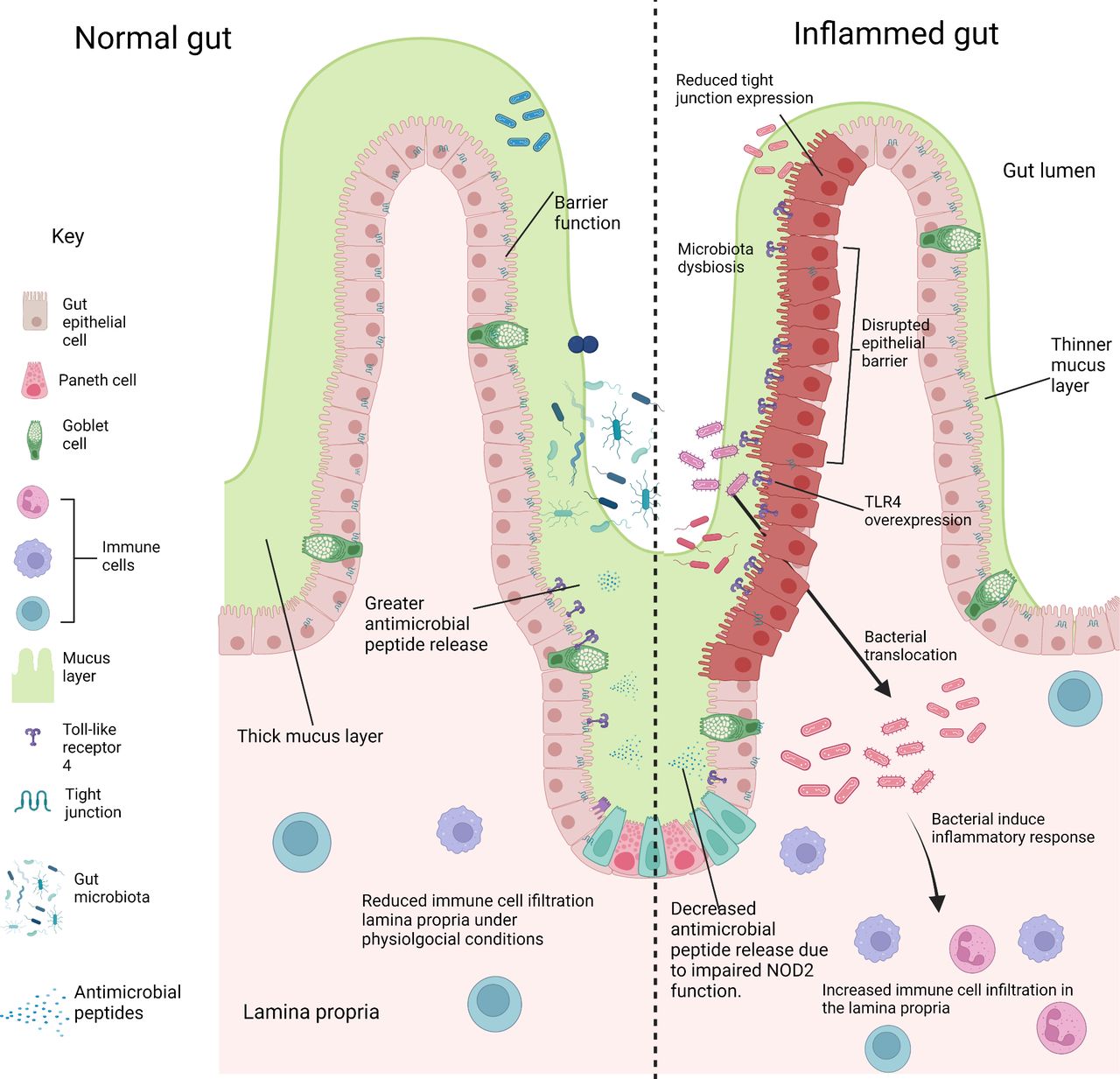
Comparing the gut environment in physiological and inflammatory conditions. Key features of a healthy normal gut are shown (left), with a thick mucus layer and intactgut epithelium preventing the entry of gut microbiota to the underlying lamina propria.Antimicrobial peptides are secreted by paneth cells into the mucus layer and gut lumen,which regulates the growth of commensals and pathogens. Disruption of these factors leads to gut inflammation (right). The mucus layer is much thinner allowing the association ofbacteria to the epithelium. Gut epithelium barrier function is impaired by the reducedexpression of tight junctions, allowing the translocation of bacteria to the lamina propria.This drives inflammation and the increased infiltration of immune cells compared tophysiological conditions. Reduced expression of antimicrobial peptides occurs due to theimpaired function of Nucleotide-binding oligomerization domain-containing protein 2(NOD2). Reduced antimicrobial peptide secretion leads to bacterial overgrowth in the gutlumen. (Figure created in BioRender).
Altered gut epithelial barrier function in disease
Another mechanism by which the microbiome might influence disease onset is by impairment of gut barrier function and resultant increase in gut barrier permeability preceding the onset of inflammation. The intestinal barrier is formed by a layer of tightly connected IECs and has several mechanisms to reduce the risk of infection by pathogenic organisms. This includes regulation of barrier permeability by tight-junctional proteins, the formation of a mucus layer, to prevent attachment of pathogens and secretion of antimicrobial peptides (figure 3).
TLR4 overexpression in the premature gut and activation by the altered microbiota in NEC negatively impacts the homoeostasis of gut epithelial cells by increasing apoptosis and reducing epithelial cell proliferation, disrupting the ability to repair the intestinal epithelium.66 84 85 A further consequence of TLR4 overexpression is the enhanced recruitment of CD4 +T cells to the gut lamina propria, as seen in mutant mice which selectively overexpress TLR4 on IECs (figure 2).86 This enhanced CD4 +T cell infiltration is also seen in the NEC mouse model for wild-type mice and in the human intestine during NEC, indicating that lymphocyte infiltration in the gut lamina propria may play an important role in NEC pathology.86 In CD, NOD2 mutations have also been linked with altered gut turnover and gut permeability such a tight junctional complex alterations.87 88
Intestinal epithelial hyperpermeability has been a well-documented feature in CD and UC, with mucosal biopsies of patients demonstrating bacterial internalisation in IECs compared with negligible mucosal bacterial load in biopsies of healthy control patients.89 Mouse models with loss of endogenous E-cadherin, which maintains the tight junctions between IECs, developed histological features of CD within 3 months.90 Similarly, in rat and human studies of NEC, there is reduced expression of both E-cadherin gene and protein with associated increase in epithelial permeability. Rho-associated protein kinase (ROCK) is involved with regulation of cadherin function, with activation of ROCK leading to disruption of apical tight junctions.91 In rat models of NEC, increased ROCK1 activation is a feature of early NEC and ROCK1 inhibition being protective in this experimental model.92
The role of the altered microbiota in initiating epithelial barrier dysfunction is still poorly understood. However, the resultant interaction between the microbiota and host clearly plays a role in driving inflammation in both diseases.
Conclusions and future directions
In this review, we have highlighted the critical role of the gut microbiota in supporting host intestinal health and have outlined two inflammatory disorders of the GIT in which the gut microbiota is involved; IBD and NEC. An altered gut microbiota is associated with these diseases and a dysregulation of the immune system is also apparent. However, whether there is a causal relationship between altered gut microbiota and the host immune response is currently unclear, although the presence of a dysfunctional epithelial barrier is an important feature in both diseases.
We have outlined possible mechanisms which begin to explain how a dysbiotic gut microbiota in conjunction with the host immune system may give rise to pathological gut inflammation. In both NEC and IBD, alterations to PRRs and their expression by IECs may be interconnected with the gut microbiota and impaired barrier function which further drives inflammation and inappropriate activation of the immune response.
Targeting these mechanistic pathways may lead to the development of novel therapies for these diseases. Small molecule TLR4 inhibitors have shown promising results, reducing severity of inflammation and disease in animal models of NEC and IBD.93 94 Probiotics, defined as ‘live microorganisms which when administered in adequate amounts confer a health benefit on the host’,95 have shown promise in reducing rates of severe NEC and mortality in preterm infants.96 97 However, evidence of their benefits remains limited, particularly for extremely low birth weight (<1000 g) infants, who are at the greatest risk of developing NEC, neither is there sufficient evidence to determine the optimal probiotic regimen.98 99 In IBD, there is limited evidence of benefit for patients with a lack of well designed and adequately powered studies.
IBD and NEC are both complex and multifactorial gut inflammatory disorders and possible disease mechanisms are only beginning to be understood. Much work has been done to characterise the gut microbiota for both diseases preceding and during disease states which has uncovered specific changes in the gut microbiota. However, whether such changes are driving the onset of such diseases is unclear and could be correlative rather than causative. A better mechanistic understanding of disease will likely arise by moving away from observational study of the gut microbiota and towards studying specific functions of member organisms, thought to be involved in disease.
Ethics statements
Patient consent for publication
References
Footnotes
DS and DE are joint first authors.
Twitter @MissLHancock, @sheencr
Contributors DS, DE and SC conceptualised and wrote the original draft manuscript. KZC, JM, AB, LH and SC reviewed and edited the original draft manuscript. SC provided oversight and overall supervision for this project.
Funding DS and DE are both PhD students at the University of Manchester with funding from the Topol Fellowship and Biotechnology and Biological Sciences Research Council (BBSRC), respectively.
Competing interests None declared.
Provenance and peer review Commissioned; externally peer reviewed.
Linked Articles
- UpFront